Dafin F. Mureșanu este profesor și șeful Departamentului de neuroștiințe la UMF „Iuliu Hațieganu” Cluj-Napoca. Este președintele Societății de Neurologie din România și al Societății pentru Studiul Neuroprotecției și Neuroplasticității. Conduce Institutul RoNeuro – Centrul de Cercetare și Diagnostic al Bolilor Neurologice
Betina Korka este cercetător la Departamentul de psihologie de la Carl von Ossietzky Universität Oldenburg
Olivia Roșu este medic rezident neurologie la Spitalul Clinic Județean de Urgență Cluj-Napoca
Dana Slovoacă este medic specialist neurolog la Institutul RoNeuro – Centrul de Cercetare și Diagnostic al Bolilor Neurologice Cluj-Napoca
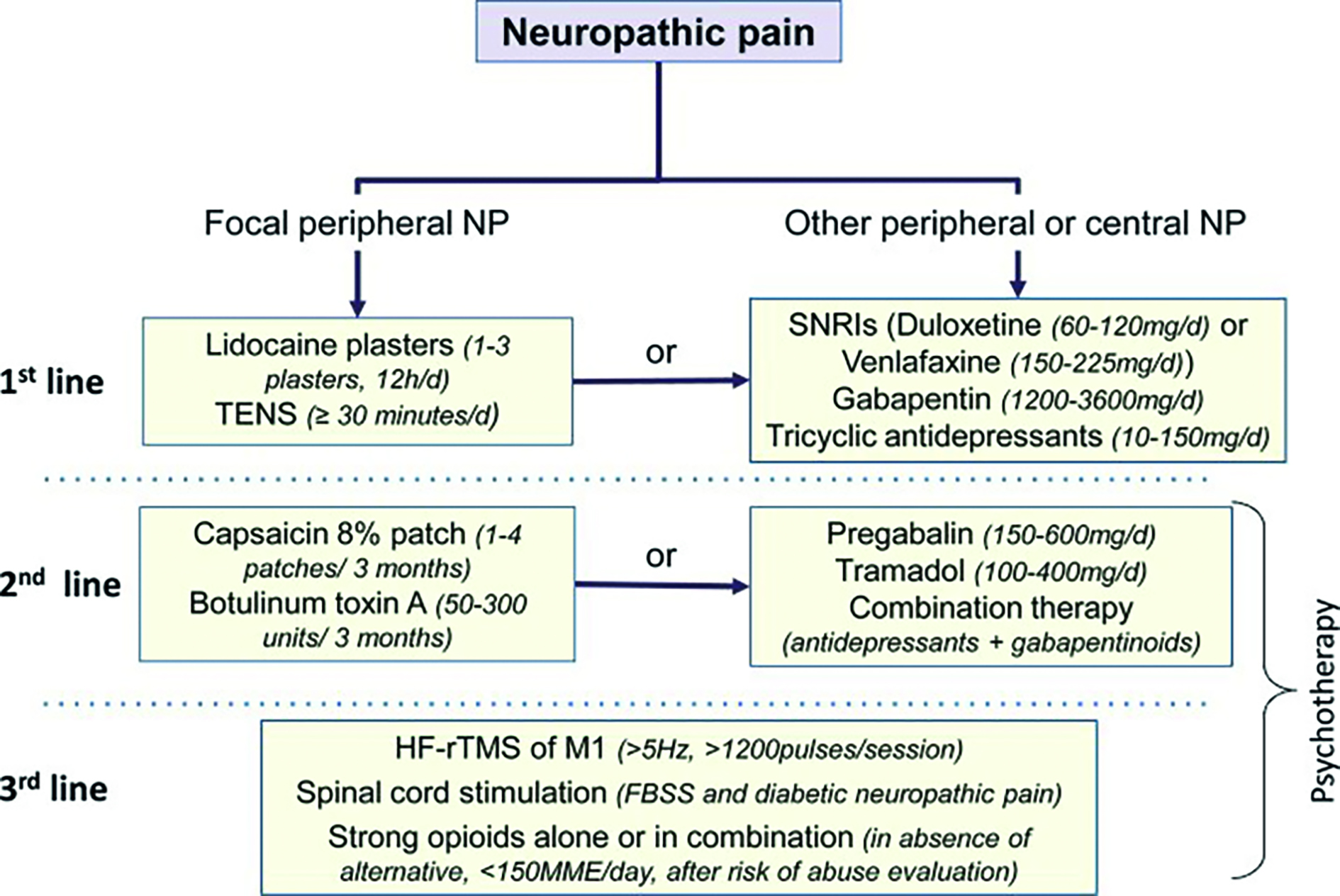
Plasticitatea patologică din durerea neuropatică este rezultatul modificărilor structurale și funcționale care reorganizează întregul sistem nervos, începând cu receptorii periferici, continuând cu sistemul nervos periferic, măduva spinării, trunchiul cerebral, cerebelul, diencefalul, corpii striați, sistemul limbic, cortexul somatosenzitiv, cortexul motor și cortexul prefrontal.
Modificări structurale
Modificări funcționale
Modificări celulare și moleculare
Modificări epigenetice
Concluzie
Prin intervenția farmacologică sau non-farmacologică la nivelul unui singur mecanism afectat, chiar dacă se obțin efecte pozitive pe moment, pe termen lung nu apare o îmbunătățire sustenabilă a procesului patologic. Sunt necesare studii preclinice și clinice care să aplice o abordare multidimensională a durerii neuropatice.
Bibliografie
1. West SJ et al. Circuitry and plasticity of the dorsal horn – toward a better understanding of neuropathic pain. Neuroscience. 2015 Aug;300:254-75
2. Tan AM, Waxman SG. Dendritic spine dysgenesis in neuropathic pain.
Neurosci Lett. 2015 Aug;601:54-60
3. Wilcox SL et al. Anatomical changes at the level of the primary synapse in neuropathic pain: evidence from the spinal trigeminal nucleus. J Neurosci. 2015;35(6):2508-15
4. Baliki MN et al. Brain Morhological Signatures for Chronic Pain. PLoS One. 2011;6(10):e26010
5. Desouza DD et al. Sensorimotor and pain modulation brain abnormalities in trigeminal neuralgia: a paroxysmal, sensory-triggered neuropathic pain. PLoS One 2013;8:e66340
6. Desouza DD et al. Abnormal trigeminal nerve microstructure and brain white matter in idiopathic trigeminal neuralgia. Pain. 2014 Jan;155(1):37-44
7. Farmer MA et al. A dynamic network perspective of chronic pain. Neurosci Lett. 2012 June 29;520(2):197-203
8. Woolf CJ. Central sensitization: implications for the diagnosis and treatment of pain. Pain. 2011 March;152(3 Suppl):S2-15
9. Apkarian AV et al. Human brain mechanisms of pain perception and regulation in health and disease. Eur J Pain. 2005;9:463-84
10. Simons L et al. The human amygdala and pain: evidence from neuroimaging. Hum Brain Mapp. 2014 February;35(2):527-38
11. Neugebauer V. The amygdala: different pains, different mechanisms. Pain. 2007 January;127(1-2):1-2
12. Lumley MA et al. Pain and emotion: a biopsychosocial review of recent research. J Clin Psychol. 2011 September;67(9):942-68
13. Windmann S et al. Role of the prefrontal cortex in attentional control over bistable vision. J Cogn Neurosci. 2006;18(3):456-71
14. Fischer TZ et al. Thalamic neuron hyperexcitability and enlarged receptive fields in the STZ model of diabetic pain. Brain Res. 2009 May 1;1268:154-61
15. Ab Aziz CB, Ahmad AH. The role of the thalamus in modulating pain. Malays J Med Sci. 2006 July;13(2):11-8
16. Nijs J et al. Applying modern pain neuroscience in clinical practice: criteria for the classification of central sensitization pain. Pain Physician. 2014;17(5):447-57
17. Youssef AM et al. Differential brain activity in subjects with painful trigeminal neuropathy and painful temporomandibular disorder. Pain. 2014 Mar;155(3):467-75
18. Liu J et al. Quantitative cerebral blood flow mapping and functional connectivity of postherpetic neuralgia pain: a perfusion fMRI study. Pain. 2013;154:110-8
19. LeBlanc BW et al. Minocycline injection in the ventral posterolateral thalamus reverses microglial reactivity and thermal hyperalgesia secondary to sciatic neuropathy.
Neurosci Lett. 2011 July;498(2):138-42
20 Loggia ML et al. Evidence for brain glial activation in chronic pain patients.
Brain. 2015 Mar;138(Pt 3):604-15
21. Tsuda M. Microglia in the spinal cord and neuropathic pain. J Diabetes Investig. 2016;7:17-26
22. Zhuo M et al. Neuronal and microglial mechanisms of neuropathic pain. Mol Brain. 2011 Jul;4:31
23. Wang D et al. Activated microglia in the spinal cord underlies diabetic neuropathic pain. Eur J Pharmacol. 2014 Apr 5;728:59-66
24. Yoo S et al. Resolvins: endogenously-generated potent painkilling substances and their therapeutic perspectives. Curr Neuropharmacol. 2013;11(6):664-76
25. Verma V et al. Nociception and role of immune system in pain.
Acta Neurol Belg. 2015 Sep;115(3):213-20
26. Luchting B et al. Anti-inflammatory T-cell shift in neuropathic pain.
Neuroinflammation. 2015 Jan 21;12(1):12
27. Gaspar JM et al. Inside the Diabetic Brain: Role of Different Players Involved in Cognitive Decline. ACS Chem Neurosci. 2016 Feb 17;7(2):131-42
28. Mizumura K, Murase S. Role of nerve growth factor in pain. Handb Exp Pharmacol. 2015;227:57-77
29. Guo W et al. Supraspinal brain-derived neurotrophic factor signaling: a novel mechanism for descending pain facilitation. J Neurosci 2006;26(1):126-37
30. Nijs J et al. Brain-derived neurotrophic factor as a driving force behind neuroplasticity in neuropathic and central sensitization pain: a new therapeutic target? Expert Opin Ther Targets. 2014 Dec;18:1-12
31. Saab CY. Pain-related changes in the brain: diagnostic and therapeutic potentials. Trends Neurosci. 2012 July;35(10):629-37
32. Descalzi G et al. Epigenetic mechanisms of chronic pain. Trends Neurosci. 2015;38(4):237-46
33. Géranton SM et al. A role for transcriptional repressor methyl-CpG-binding protein 2 and plasticity-related gene serum-and glucocorticoid-inducible kinase 1 in the induction of inflammatory pain states. J Neurosci. 2007; 27(23):6163-73
34. Géranton SM et al. Descending serotonergic controls regulate inflammation-induced mechanical sensitivity and methyl-CpG-binding protein 2 phosphorylation in the rat superficial dorsal horn. Mol Pain. 2008 Sep;4:35
35. Kynast KL et al. Modulation of central nervous system-specific microRNA-124a alters the inflammatory response in the formalin test in mice. Pain. 2013;154(3):368-76
36. Viet CT et al. Re-expression of the methylated EDNRB gene in oral squamous cell carcinoma attenuates cancer-induced pain. Pain. 2011;152(10):2323-32
37. Bienvenu T, Chelly J. Molecular genetics of Rett syndrome: when DNA methylation goes unrecognized. Nat Rev Genet. 2006;7(6):415-26
38. Fuks F et al. The methyl-CpG-binding protein MeCP2 links DNA methylation to histone methylation. J Biol Chem. 2003;278(6):4035-40
39. Imai S et al. Change in micro-RNAs associated with neuronal adaptive responses in the nucleus accumbens under neuropathic pain. J Neurosci. 2011;31(43):15294-9
40. Chiechio S et al. Epigenetic modulation of mGlu2 receptors by histone deacetylase inhibitors in the treatment of inflammatory pain. Mol Pharmacol. 2009;75(5):1014-20
Dacă vrei să fii la curent cu tot ce se întâmplă în lumea medicală, abonează-te la „Viața Medicală”, publicația profesională, socială și culturală a profesioniștilor în Sănătate din România!
Titularii abonamentelor pe 12 luni sunt creditați astfel de:

Dacă vrei să fii la curent cu tot ce se întâmplă în lumea medicală, abonează-te la „Viața Medicală”, publicația profesională, socială și culturală a profesioniștilor în Sănătate din România!
Află mai multe informații despre oferta de abonare.
Cookie-urile ne ajută să vă îmbunătățim experiența pe site-ul nostru. Prin continuarea navigării pe site-ul www.viata-medicala.ro, veți accepta implicit folosirea de cookie-uri pe parcursul vizitei dumneavoastră.
Da, sunt de acord Aflați mai multe