Dezvoltarea actuală a
neuroştiinţelor a condus la argumente promiţătoare în legătură cu recuperarea
funcţiilor cognitive în afecţiunile nevraxiale şi, mai ales, în afecţiunile
vasculare. Activarea neuronală şi cunoaşterea mai precisă a apoptozei sunt cele
două modalităţi prin care, în prezent, se pot aplica modalităţi de intervenţie
nu numai în suferinţa neuronului periferic, ci şi în neuronul central, unde
sistemul glial conduce la un nou mod de interpretare a mecanismelor bolii, în
sensul obţinerii unor rezultate favorabile de ameliorare metabolică şi,
respectiv, funcţională în urma suferinţelor cerebrale vasculare sau non-vasculare.
Cea mai mare parte din operaţiile realizate
de creier depind de interconexiunile foarte precise dintre cele 100 de miliarde
de neuroni. Organizarea sistemului vizual, de la retină, corpul geniculat
lateral, până la cortex ilustrează importanţa acestui mecanism. Reţelele
neuronale sunt elaborate având ca punct de plecare instrucţiunile genetice care
permit axonilor detectarea corectă a traiectului lor, atingând ţintele exacte.
Totuşi, realizarea definitivă a circuitelor nervoase depinde în mare măsură de
informaţiile senzoriale care vin din mediul exterior, în special în perioada
copilăriei.
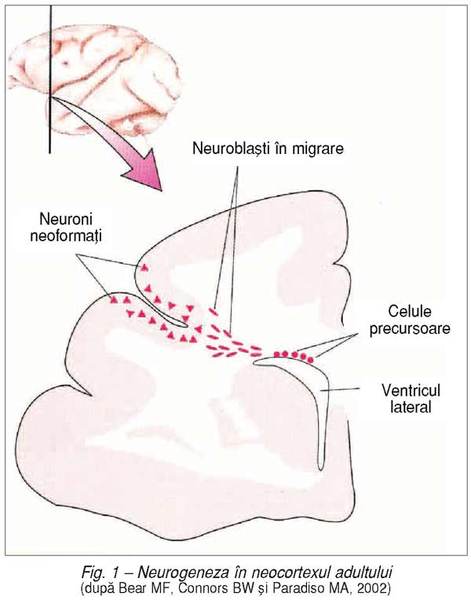
Originea
neuronilor. Prima fază de organizare a circuitelor sistemului nervos priveşte
originea neuronilor. Dezvoltarea structurii neuronale cunoaşte trei faze principale:
proliferarea, migrarea şi, respectiv, diferenţierea celulară.
Geneza
conexiunilor neuronale (fig. 1). În
cursul diferenţierii lor, neuronii emit axoni, care vor întâlni ţintele cele
mai apropiate. Se poate considera că dezvoltarea acestei vaste reţele de
conexiuni sau formarea de căi se desfăşoară în trei faze: selecţia traiectului,
selecţia ţintei şi selecţia destinaţiei. Acest lucru a fost bine demonstrat în
contextul dezvoltării căilor vizuale, care pornesc de la retină la corpul
geniculat extern. Când neuroblastul, în migrarea sa, găseşte destinaţia adecvată
în sistemul nervos, se diferenţiază şi neuronul emite prelungiri, care formează
axonul şi dendritele. În această fază precoce, totuşi, prelungirile axonale şi
dendritice sunt foarte asemănătoare şi sunt calificate drept neurite.
Extremitatea în creştere a unei asemenea structuri este aşa-zisul con de creştere. Acesta serveşte la
recunoaşterea traiectului de către structurile respective. Extremitatea
exploratoare a conului de creştere este compusă din straturi membranare
aplatizate, lamelipode, care
ondulează în mişcarea de finalizare a procesului de creştere. Alte structuri – filopodele – se întind şi se retractă în
mod constant, explorând mediul înconjurător.
Elongaţia axonilor de-a lungul unei „rute moleculare“ este facilitată de procesele defasciculaţie, mecanism care
contribuie la asocierea între axonii care se dezvoltă în ansamblu. Fasciculaţia
este expresia moleculelor de suprafaţă, denumite molecule de adeziune (sau CAMs
– cell adhesion molecules). CAMs de la nivelul membranelor axonale vecine
interacţionează imediat, contribuind la o creştere a tuturor axonilor în acelaşi
timp.
O altă problemă se referă la ghidajul axonal. Realizarea căilor
neuronale reprezintă o preocupare excepţională din punct de vedere ştiinţific,
dacă avem în vedere distanţele foarte importante pe care anumiţi axoni trebuie
să le parcurgă în creierul adultului. Orientarea către ţinte se realizează şi
cu ajutorul unor molecule de ghidaj, care pot avea caracter atractiv sau
repulsiv, în funcţie de natura moleculelor şi de receptorii prezenţi la nivelul
axonilor aflaţi în dezvoltare. De asemenea, un factor chemoatractiv este şi molecula difuzibilă care acţionează la distanţă
pentru a „atrage“ axonii în dezvoltare către ţinta proprie. Existenţa acestor
factori a fost propusă de aproape un
secol de Ramón y Cajal, dar numai foarte recent aceşti factori au fost
identificaţi. Unul dintre ei este netrine,secretat de neuroni la nivelul părţii ventrale şi mediane a măduvei spinării.
Formarea
sinapselor are
loc atunci când conul de creştere intră în contact cu ţinta sa. El se
aplatizează şi formează o sinapsă. Interacţiunea între axon şi ţintă are un
dublu sens şi inducţia terminaţiei la nivelul presinaptic implică, de asemenea,
acţiunea proteinelor de la nivelul lamei bazale. Factorii legaţi de lama bazală
aduşi prin intermediul celulelor ţintă pot stimula intrarea calciului în conul
de creştere, declanşând astfel eliberarea de neurotransmiţători.
Eliminarea
celulelor şi sinapselor. Într-o perioadă lungă de dezvoltare, de la naştere
până la vârste înaintate, aceste conexiuni sunt supuse unor procese de reducere
drastică a numărului de neuroni şi sinapse. Este vorba de moartea celulară cunoscută sub numele de moarte celulară programată, un proces care se accentuează în timp.
Când axonii şi-au atins ţintele şi s-au format structurile sinaptice, numărul
de axoni presinaptici şi de neuroni scade progresiv. Moartea celulară este în
relaţie cu o competiţie faţă de factorii
trofici, adică substratul nutritiv necesar supravieţuirii ţesuturilor, ca şi
celulelor ţintă, furnizate în cantitate limitată.
Alături de moartea celulară, apar modificări ale capacităţilor sinaptice. În acest caz, fiecare neuron primeşte un
număr determinat de sinapse asupra dendritelor sale, precum şi asupra somei.
Aceasta reprezintă capacitatea sinaptică
a neuronului. În toate structurile nevraxiale, în primele faze de
dezvoltare, capacitatea sinaptică este puternică şi scade când neuronul ajunge
la maturitate. Modificările capacităţii sinaptice neuronale în cursul dezvoltării
creierului se observă cel mai bine la joncţiunea neuromusculară. Într-un prim
stadiu, o fibră musculară poate să primească aferenţe pornind de la mai mulţi
motoneuroni. Totuşi, această inervaţie polineuronală dispare şi fiecare fibră
musculară primeşte în final aferenţe sinaptice de la un singur motoneuron alfa.
Reorganizarea
sinaptică depinde de activitate (fig. 2). Aceste modificări
trebuie evaluate în procesul de recuperare. Reorganizarea sinaptică reprezintă
etapa finală în procesul de selecţie a destinaţiei. În sens invers, o serie de
etape în faze iniţiale ale formării conexiunilor neuronale arată că, totuşi, reorganizarea sinaptică este o consecinţă a
activităţii neuronale şi a transmisiei sinaptice.
David Hubel şi Torsten Wiesel, studiind
sistemul vizual, au arătat capacitatea de dezvoltare a căilor compensatorii,
fapt ce a condus la cercetări promiţătoare în alte structuri neuronale
implicate în procesul de recuperare funcţională. Este util să evocăm aşa-zisul
concept de „perioadă critică“.
Aceasta se referă la existenţa unui moment când evoluţia celulelor este
susceptibilă de a fi modificată de interacţiunile celulare. Acest concept a
fost dezvoltat de embriologul Hans Spemann, care a arătat că transplantarea
unei părţi dintr-un embrion la altul trebuie să se facă într-o anumită perioadă
bine precizată a dezvoltării, deoarece mecanismele de interacţiune celulară pot
afecta fenotipul celulelor transplantate. Atunci când un muşchi nu funcţionează,
apare atrofia şi se pierde forţa musculară. Este vorba de eliminarea
sinapselor, private de activitate. La nivelul cortexului striat, acest fenomen
pare să nu se producă, căci modificările în raport cu informaţiile care ajung
la nivelul cortexului pot fi într-un proces de competiţie. Competiţia care se stabileşte la nivelul cortexului
vizual, de exemplu, este demonstrată de consecinţele unui strabism. Este de
subliniat un fapt clinic esenţial şi anume: cataractele congenitale, de
exemplu, sau lipsa de paralelism ocular trebuie să fie corijate precoce, pentru
a evita un handicap vizual definitiv. Plecând de la acest exemplu, se poate
discuta de aşa-zisa influenţă modulatorie,
care are loc în evoluţia structurală şi care poate fi activată de circuite
neuronale corticale în raport cu stimulii adecvaţi.
Mecanismele
elementare ale plasticităţii sinaptice corticale. Două principii de bază
pot fi avansate în ceea ce priveşte modificările sinaptice: 1) când neuronul
presinaptic este activ şi, în acelaşi timp, neuronul postsinaptic este puternic activat sub influenţa altor
aferenţe nervoase, atunci sinapsa formată de neuronul presinaptic este activată.
Aceasta este o altă modalitate de formulare a teoriei lui Hebb. În alţi
termeni, neuronii care descarcă în
ansamblu se dezvoltă în ansamblu; 2) Când neuronul presinaptic este activ şi,
în acelaşi timp, neuronul postsinaptic nu este decât slab activat de alte aferenţe nervoase, atunci sinapsa formată de
neuronul presinaptic va reacţiona mai slab. Deci
neuronii care nu prezintă sincronism în procesul de activare prin descărcare
conduc la o scădere a activităţii funcţionale.
Se impune deci corelaţia. Pentru a obţine
o activare „extinsă“, activitatea sinaptică trebuie să fie corelată cu cea
legată de numeroase alte aferenţe sinaptice care converg către acelaşi neuron
postsinaptic. Atunci când activitatea sinaptică este corelată cu o activare
persistentă având un puternic răspuns postsinaptic, sinapsa este selecţionată şi
activitatea sa întărită. În caz contrar, atunci când activitatea unei sinapse
nu este corelată cu neuronul postsinaptic, sinapsa slăbeşte în activitate şi
este eliminată. Din acest punct de vedere, sinapsele sunt „validate“ pe baza
capacităţii lor de a participa eficace la descărcările partenerului
postsinaptic.
Care sunt mecanismele responsabile de
asemenea modificări care se bazează pe corelarea descărcărilor sinaptice? Răspunsul
se află la nivelul mecanismelor de transmitere excitatorii ale sistemului
nervos central (fig. 3).
Se cunoaşte că mecanismele plasticităţii
sinaptice susceptibile de a explica fenomenele de învăţare sunt, în fapt,
comparabile cu mecanismele cunoscute de reorganizare sinaptică în cursul
dezvoltării. Există o serie de perioade critice mai puţin cunoscute în aceste
procese. Se pot dezvolta trei ipoteze: 1)
Plasticitatea se diminuează atunci când creşterea axonului încetează; 2)
Plasticitatea se diminuează atunci când transmisia sinaptică se află în faza
completă de maturitate. Se cunoaşte proprietatea receptorilor NMDA de a se
modifica în cursul perioadei critice; 3) Plasticitatea se diminuează atunci când
activitatea corticală este inhibată.
Întrebarea cu privire la sfârşitul
perioadelor critice este importantă. Modificările sinaptice şi reorganizările
anatomice ale circuitelor permit o anumită recuperare funcţională atunci când
sistemul nervos este afectat. Totuşi, recuperarea este, din păcate, foarte
limitată în creierul adultului. Recuperarea funcţională după o leziune a
creierului poate fi cvasitotală în sistemul nervos la tineri, în cursul dezvoltării,
atunci când reorganizarea sinaptică este încă posibilă. De asemenea, una din
ideile importante se referă la înţelegerea mecanismelor plasticităţii din
cursul dezvoltării normale, ce se pot adapta la mijloacele de favorizare a unei
recuperări funcţionale tardive, în cazurile de leziuni vasculare cerebrale de
exemplu.
Neuroplasticitatea, învăţarea şi memoria
Formarea conexiunilor neuronale reprezintă
un proces extraordinar de bine organizat, ce urmează anumite reguli bine
precizate. De la naştere (şi, fără îndoială, şi înainte de naştere), stimulii
senzoriali modifică funcţionarea şi organizarea creierului şi influenţează
comportamentul.
Dezvoltarea structurilor cerebrale asociate
experienţei este strâns legată de învăţare. În această perspectivă, învăţarea şi
memoria pot fi considerate drept adaptări permanente ale circuitelor cerebrale
la mediu, dând posibilitatea unui răspuns corect la diferite situaţii.
Diferitele
tipuri de memorie şi de amnezie. Procesul de învăţare este achiziţia de noi informaţii sau cunoştinţe şi memoria corespunde reţinerii informaţiilor
dobândite. În toată existenţa umană se învaţă şi memorează numeroase date cu
caracter diferit şi trebuie subliniat că toate aceste lucruri nu sunt tratate şi
stocate de aceleaşi procese neuronale. Deci, în procesul de învăţare/recuperare,
învăţarea nu implică o singură structură cerebrală sau un singur mecanism
celular. În plus, procesele de stocaj al unor informaţii particulare pot varia în
fiecare moment.
Memoria
declarativă şi memoria non-declarativă. În general, memoria declarativă este
disponibilă pentru o reamintire conştientă, ceea ce nu este cazul pentru
memoria non-declarativă. Memoria declarativă corespunde, în general, la ceea ce
numim „amintire“ în limbajul curent, în timp ce memoria procedurală este mai
apropiată de „obişnuinţă“ sau de obişnuinţa dobândită.
Memoria
pe termen scurt şi memoria de lungă durată se referă la amintiri de zile,
luni sau ani după achiziţie.
Memoria de
scurtă durată
este de ordinul câtorva secunde sau ore şi este relativ labilă. Consolidarea mnezică este procesul
necesar şi posibil de realizat.
Amnezia
În viaţa cotidiană, uitarea este un fapt la
fel de obişnuit ca şi învăţarea. Acest proces este normal şi inevitabil. Totuşi,
diferitele leziuni cerebrale, în special vasculare, dar şi non-vasculare, pot
interfera cu procesele mnezice.
În cazul unei amnezii date de aceste
leziuni, amnezia retrogradă se
manifestă prin uitarea evenimentelor ce s-au derulat în timpul perioadei care a
precedat leziunea, dar amintirile mai vechi sunt prezervate. În cazul amneziei anterograde, evenimentele care
au precedat traumatismul, de exemplu, sunt conservate, dar subiectul nu este
capabil de a reda amintirile ce au avut loc după traumatism (fig. 4).
Amnezia globală
tranzitorie este
o altă formă de amnezie, care nu durează decât puţin timp. Ea apare brutal, ca
un episod de amnezie anterogradă ce nu durează decât câteva minute sau câteva
zile, asociată cu o amnezie retrogradă pentru evenimentele recente care au
precedat acest episod. Acest tip de amnezie este în general legat de o scurtă
ischemie cerebrală, când debitul sanguin este temporar redus la nivelul creierului. Cazurile de amnezie globală tranzitorie – asociate stresului excesiv,
folosirii drogurilor şi duşurilor reci – au fost raportate, întrucât aceste
evenimente afectează debitul sanguin cerebral.
creierului. Cazurile de amnezie globală tranzitorie – asociate stresului excesiv,
folosirii drogurilor şi duşurilor reci – au fost raportate, întrucât aceste
evenimente afectează debitul sanguin cerebral.
Cercetarea engramelor
Reprezentarea fizică sau localizarea
memoriei este denumită engramă.
Cunoaşterea engramelor a beneficiat de metode experimentale numeroase, metoda
labirintului fiind cea mai cunoscută. Teoria
lui Hebb (fig. 5) şi teoria ansamblului celular arată că
ideile lui Hebb au favorizat dezvoltarea unor modele matematice ale circuitelor
neuronale, conducând la rezultate promiţătoare în procesele de influenţare a
fenomenelor de stocaj mnezic.
După teoria lui Hebb, dacă o engramă se
bazează pe informaţia transmisă printr-o singură modalitate senzorială, va
trebui să permită localizarea zonelor cortexului corespunzătoare acestui
sistem.
Lobul temporal şi memoria declarativă
Asocierea lobului temporal este probată în
ceea ce priveşte rolul particular de stocare a evenimentelor trecute. Lobii
temporali sunt formaţi de cortexul temporal, care este un loc de stocaj al memoriei
de lungă durată. Trebuie menţionat că în această structură se găsesc hipocampul
şi alte structuri, esenţiale în formarea amintirilor memoriei declarative.
Hipocampul este o structură situată în raport cu ventriculul lateral. Trei
regiuni importante situate ventral de hipocamp – cortexul entorinal, cortexul peririnal şi cortexul parahipocampic – participă la procesul de memorie. Una din căile eferente majore ale
hipocampului este fornixul, care descrie o buclă în jurul talamusului, ajungând
la hipotalamus.
În procesul de memorie, diencefalul reprezintă o zonă extrem de
sensibilă în procesele mnezice. Leziunile vasculare cerebrale de la nivelul
cortexului cerebral şi, desigur, cele cu localizare talamo-hipotalamică, sunt întâlnite
frecvent şi cu numeroase particularităţi la nivelul perturbărilor microcirculaţiei.
O serie de cercetări au arătat că striatum reprezintă o structură critică
în ceea ce priveşte memoria procedurală implicată
în formarea abilităţilor motorii.
În esenţă, învăţarea şi memoria nu pot fi
localizate doar într-o singură regiune a creierului. Nu putem vorbi de un număr
de celule specializate sau de „celule pentru memorie“, a căror activitate serveşte
stocării informaţiilor legate de experienţa de viaţă, fără niciun raport cu
alte funcţii ale creierului.
Problema care se pune se referă la înţelegerea
modului în care interacţionează toate aceste regiuni cerebrale pentru a ne
permite înţelegerea. Se poate spune, totuşi, că memoria de lungă durată prezintă
o formă mai puţin labilă şi rezistă întreruperilor la anestezie, ca şi în cazul
unor traumatisme inerente. În fapt, organizarea creierului se poate modifica
constant, pentru a se adapta experienţelor vieţii, fapt ce stă la baza
mecanismelor de învăţare şi memorizare.
Mecanismele
moleculare de învăţare şi de memorie
Este vorba, în primul rând, de
plasticitatea sinaptică a cortexului cerebelos, a hipocampului şi a
neocortexului, precum şi de bazele moleculare ale memoriei de lungă durată, în
care sinteza proteinelor şi consolidarea mnezică conduc la aşa-zisa
plasticitate structurală şi memorie.
Sinapsa poate fi locul mecanismelor de învăţare
şi de memorie. Indiferent de locul în care se află aceste sinapse şi de tipul
de memorie, un mare număr de mecanisme subiacente par să fie universale.
Manifestarea evenimentelor care se desfăşoară se traduce, în primul rând, prin
modificări ale activităţii electrice cerebrale, în al doilea rând prin intervenţia
de mesager molecular şi, în final, prin modificări ale proteinelor sinaptice
preexistente. Aceste modificări ale caracterului tranzitoriu sunt convertite în
modificări durabile – memorie de lungă durată – prin schimbarea structurală a
sinapsei însăşi. În numeroase modele de studiu cu privire la memorie, aceste
procese implică sinteza unei noi proteine şi organizarea de noi microcircuite. În
toate cazurile, procesul de învăţare este asociat unor numeroase mecanisme ce
sunt utilizate pentru declanşarea circuitelor neuronale în curs de organizare, în
timpul procesului de învăţare. Unul din aspectele comune în toate aceste modele
este participarea ionilor de Ca2+. Ionul de calciu nu serveşte doar la
consolidarea osului, nu joacă numai un rol critic în secreţia de neurotransmiţători
şi în contracţia musculară, ci este, de asemenea, unul din factorii de
plasticitate sinaptică, un puternic mesager. Ca2+ are capacitatea unică de a
cupla în mod direct activitatea electrică cu modificările de lungă durată de la
nivelul creierului.
Plasticitatea sinaptică a hipocampului şi a
neocortexului, pe de o parte, şi a structurilor cerebeloase, pe de altă parte,
potenţează şi modelează activităţile cognitive în raport cu timpul.
În
concluzie, datele de mai sus reprezintă elementele ştiinţifice de bază cu
privire la neuroplasticitate ce se studiază în etapa actuală de cunoaştere.
Fiecare din ideile principale – sintetizate
extrem de succint – ne demonstrează substratul neurobiologic, celular şi
molecular, atât în procesul de dezvoltare, cât şi în genetica apoptozei, care
se accentuează odată cu vârsta înaintată.
În fiecare etapă a dezvoltării fenotipice,
neuroştiinţele sugerează, la nivel celular (şi, în acelaşi timp, global),
numeroase posibilităţi de reversibilitate a unor perturbări neuronale, mai ales
sinaptice, precum şi eventuale riscuri de intervenţii neadecvate faţă de un
proces natural de neuroplasticitate şi recuperare funcţională.
Substratul metabolic al acestor modificări
care încep în viaţa embrionară şi până la vârstele cele mai avansate este astăzi
realizat de substanţa albă şi de alte structuri – întreaga structură glială,
prin intermediul microcirculaţiei cerebrale, într-o complexitate funcţională
care este în faza de descifrare la om, bazându-ne, din păcate, numai pe datele
experimentale.
Patologia vasculară cerebrală nu cuprinde
numai entităţile clasice bine cunoscute, ci şi modificări vasculare de cele mai
multe ori fără expresie clinică, dar care necesită o cunoaştere adecvată,
deoarece intervenţia terapeutică potrivită este dictată de modul în care
oxigenul şi glucoza ajung să fie utilizate acolo unde putem vorbi de o activare
neuronală şi de o conservare a unui mecanism natural de adaptare.
În metodele de recuperare specifică
neurologică după stroke este nevoie de o cunoaştere cât mai riguroasă a tuturor
mecanismelor care suferă în procesele cognitive descrise mai sus şi aceasta
este premisa unor rezultate favorabile în procesul de recuperare. Mijloacele
fizice sau/şi medicamentoase trebuie să ţină seama de această cerinţă esenţială
şi credem că, în viitor, vom putea să obiectivizăm aceste mecanisme.
Procesul de recuperare în patologia
vasculară trebuie activat în permanenţă şi nu se opreşte niciodată, dacă se
observă constant un anumit progres.
Neuroplasticitatea în raport cu suferinţa
vasculară ne oferă argumente promiţătoare în legătură cu atenuarea
conservatorismului care a dominat până în ultimele decenii, cu privire la
imposibilitatea obţinerii unei recuperări funcţionale în procesele lezionale
cerebrale, care se referă la complexul sinaptic extraordinar realizat de cei o
sută de miliarde de neuroni, înconjuraţi de încă zece miliarde celule gliale
din substanţa albă.
Boala vasculară cerebrală trebuie să fie
considerată în permanenţă o boală generală. „Creierul vascular“ cuprinde
ansamblul morfologic şi funcţional în condiţii normale şi patologice, aflându-se
într-un continuu proces de cunoaştere, în scopul prevenirii şi tratării bolii
vasculare cerebrale.